I GENI RACCONTANO LA NOSTRA STORIA
Antropologia molecolare e genetica svelano il percorso di
Homo Sapiens
Dr. Francesco Belli
Docente di Immunologia Università “La Sapienza”, Roma
Corso di Laurea in Biotecnologie
Il percorso culturale che ha portato all' applicazione delle nuove tecniche molecolari nello studio del passato umano.
Una delle acquisizioni raggiunte dal raffronto fra antichi e moderni genomi nell'ambito della nostra e di altre specie viventi è la sostanziale identità evolutiva, considerate le caratteristiche molecolari, fra individui odierni e popolazioni di un passato antico o più recente, riemersa dalla documentazione fossile mediante tecnologie innovative. Gli sviluppi di queste ultime stanno rivoluzionando le conoscenze, nelle popolazioni umane, animali e vegetali, della storia evolutiva, i cambiamenti anatomici e fisiologici, l' impatto con le patologie. Le nuove discipline contribuiranno ad un'interazione fra tecnologie applicate, dottrine naturalistiche e dati archeologico-storici, finora spesso disattesa: l' antropologia molecolare può dare alla ricostruzione della storia evolutiva dell'uomo un contributo decisivo.
E’ tra 1700 e 1800 che la scienza inizia ad occuparsi del passato dell’uomo e soprattutto a comprendere che le nostre radici vanno ricercate in un’epoca assai più lontana di quanto credenze, fonti scritte e tradizioni religiose sulle origini avevano fino ad allora delineato. In precedenza, troviamo sporadici accenni al problema delle origini dell’umanità a margine di alcune pagine dei codici di Leonardo, il quale per primo comprese la natura dei fossili e fissò nel “diluvio universale” uno spartiacque nella storia del mondo; ulteriori cenni, naturalmente controcorrente al pensiero ufficiale, sono rintracciabili negli scritti di due eretici: Bruno e Galilei. In effetti il tema del diluvio universale ricorre in numerose culture orientali ed occidentali: in quella giudaico-cristiana riconduceva in ambito biblico l’origine di tutti i popoli, come narra la genesi: la storia inizia dopo il diluvio.
Le discipline che si andarono affermando come riferimenti su base scientifica per lo studio del nostro passato furono archeologia e antropologia; la prima iniziò a svilupparsi scientificamente alla fine del XVIII secolo, sullo spirito dell’ “Encyclopedie” e del neoclassicismo, la seconda nel corso dell’ 800, grazie alla convergenza di più fattori: il darwinismo, il positivismo, le ultime esplorazioni, il ritrovamento dei primi fossili di ominidi.
Tra XVIII e XIX secolo nasce l’ archeologia come disciplina scientifica, dapprima elitaria, poi diffusa e popolare. Ricordiamo J.J.Winckelmann (1717-68), ritenuto il fondatore dell’archeologia classica, più come teorico che come operatore sul campo e limitatamente al mondo greco-romano, che anzi considerava le civiltà egizia e del vicino oriente “inferiori”, con convinzione discriminatoria; la campagna napoleonica in Egitto (1799-1801), che al di là delle considerazioni storiche e dei successi scientifici, fu un evento di propaganda e pro-regime: la scienza al servizio dei potenti; infine H.Schliemann che convertì l’archeologia da scienza d’elite in materia popolare e diffusa: diede corpo e consistenza a miti ed eroi della tradizione letteraria classica occidentale.
Ma l’archeologia non si limita a disseppellire manufatti, costruzioni, oggetti preziosi: tornano nel mondo dei vivi fossili millenari oppure ossa di un passato recente. Pensiamo al lavoro di Schliemann che attribuì agli scheletri micenei o troiani i nomi dell’epica omerica, fallendo di alcuni secoli l’esatta datazione storica di quei reperti. L’interpretazione di uno scavo archeologico richiede sempre un intervento antropologico, tanto più quanto lontani da noi, nel tempo e nella cultura che riemerge, riappaiono uomini e oggetti; il loro insieme, la loro collocazione ragionata raramente si prestano ad una facile lettura, se non con l’apporto multidisciplinare di specialisti di aree diverse.
Nel XIX secolo, tra Darwin, positivismo, progressi scientifici e colonialismo, assistiamo pertanto allo sviluppo dell' antropologia fisica e dell' antropologia culturale. Va senz'altro menzionata la controversa figura di J.F. Blumenbach, esaltato come il padre dell’ antropologia fisica, denigrato come il fondatore del razzismo scientifico a seguito della sua classificazione della specie umana in cinque razze in base a criteri fisici evidenti come il colore della pelle: caucasica, etiopica, africana, malese, americana. In realtà egli parlò di un’unica specie umana di cui esistono cinque varietà, ma ebbe il torto, sottolineato e ripreso dai suoi epigoni, di identificare in alcuni caucasici, i georgiani in particolare, gli uomini più belli (anche perchè bianchi), da cui l’umanità intera deriva. Da lui nasce senz’altro l’antropometria, impiegata per oltre cent’anni per differenziare singoli o gruppi di uomini. Antropologia culturale, etnografia, demografia e altre discipline umanistiche si diffondono con le opere di A. De Gubernatis e i suoi studi sull’universo antico e attuale indiano e J. Frazer, che si riteneva discepolo di Darwin, attento a riti, magie e usanze dei “selvaggi”, di vita e di morte. Molte di queste opere ci appaiono oggi abbastanza ingenue e frutto di Autori che viaggiavano poco o nulla tra i popoli e le terre che descrivevano, lontani da un’esperienza sul campo che antropologi ed etnologi considereranno indispensabile solo nel corso del ‘900.
Ma è anche l'epoca in cui si impongono ideologie razziste, su presunte basi “scientifiche": numerose le opere di antropologi, medici e di altre discipline che, direttamente o volutamente distorte, hanno contribuito al diffondersi delle ideologie sulla supremazia di una razza (solitamente quella bianca) sulle altre, in un secolo di trionfante colonialismo e sfruttamento economico e sociale di interi continenti. Ricordiamo gli scritti del già citato Blumenbach, di J.A. de Gobineau, dall’inequivocabile titolo “saggio sull’ineguaglianza delle razze umane” e di H.S.Chamberlain che fu uno dei principali ispiratori ideologici dell’arianesimo, della sua supremazia e della necessità di eliminare inferiori come gli ebrei e disabili perché contaminanti.
Il 1856 è ricordato come una data epocale per la paleoantropologia e la cultura europea: nel cuore della Germania, nella valle di Neander, emersero casualmente fossili di un ominide che sarà poi denominato, insieme a molti altri ritrovati in seguito in Europa ed Asia ,“h. neanderthalensis”. Fu soprattutto uno shock culturale: in piena epoca colonialista, ove si riaffermava la supremazia dei bianchi su ogni altra etnia, nel cuore dell’ Europa in cui prendevano sempre più corpo ideologie razziste, il ritrovamento di reperti di antichi ominidi, con caratteri anatomici in parte a noi simili, ma anche “grossolanamente” diversi, pose questioni interpretative e tassonomiche per quei tempi irrisolvibili. Da qui una serie di errori clamorosi da parte della scienza ufficiale: R.Virchow affermò che i fossili dei Neandertaliani non potevano essere di un “umano normale”, ma affetto da gravi patologie ossee. Solo di recente l’ antropologia molecolare ha dimostrato che h.sapiens e h.neanderthalensis sono due specie distinte, pur avendo un antenato ancestrale comune. Anche se entrò in conflitto con le ideologie razziste del nazionalsocialismo, sui testi di Virchow si formarono numerosi medici che prestarono servizio nei lager. C.Lombroso inserì il cranio di un h. neanderthalensis tra i folli e i delinquenti; inoltre sostenne sempre l'esistenza di differenze fisiche e mentali fra italiani del nord e meridionali. In tempi recenti F.Crick, uno dei padri della doppia elica del DNA, ha "dimostrato" l’inferiorità Intellettuale e cognitiva dei negri.
Oggi antropologia molecolare e genetica hanno dimostrato in maniera definitiva che la classificazione e la suddivisione della specie umana in razze è scientificamente inconsistente ed hanno decostruito ogni possibile tentativo di farlo. La nostra specie è universalmente unica pur in una complessità variegata: come già aveva notato Darwin, “le differenze fra le popolazioni, anche se vistose, sono perlopiù irrilevanti, mentre vi è una grande uniformità nelle caratteristiche veramente importanti, comprese quelle mentali: nonostante le differenze esteriori tra aborigeni americani, negri africani ed europei quello che veramente colpisce, dai tanti aspetti del carattere, è quanto le loro menti sono simili tra loro”. L'antropologo ed etnologo C.Levi-Strauss e il genetista L.L.Cavalli-Sforza hanno scritto libri fondamentali che bocciano scientificamente l'esistenza delle razze. R.Levi-Montalcini ha affermato: "La continua variazione della specie umana è stata plasmata dagli incessanti spostamenti e dalle relazioni con gli ecosistemi".
Note tecniche.
L’antropologia molecolare è una nuova disciplina che in questi ultimi anni ha avuto un enorme sviluppo grazie a tecnologie innovative ed applicazioni biomolecolari che permettono analisi sempre più approfondite, in tempi rapidi e a costi relativamente contenuti del DNA antico (aDNA) e moderno. Dall’incontro fra antropologia molecolare e paleontologia nascono nuove opportunità di studio su antichi genomi di ominidi, animali e piante. In particolare, l’analisi di aDNA umano permette la ricostruzione: della storia naturale e dell’evoluzione della nostra specie, delle relazioni fra h. sapiens ed altre specie ominine estinte, del quadro storico e geografico delle migrazioni umane e le relazione con altri marcatori di evoluzione: ad es, le lingue; la costruzione di orologi molecolari ed alberi filogenetici, il confronto fra dati molecolari, archeologici e antropologici “classici”.
aDNA è definito come il residuo di materiale genetico che si può estrarre da numerosi campioni biologici6, d’età e stato di conservazione differente. Il caso più frequente è quello del DNA recuperabile da ossa antiche e fossili, proveniente dagli osteociti che vanno incontro ad autolisi post-mortem, i cui acidi nucleici rimangono in parte isolati e protetti in nicchie e lacune nell’ambito della matrice interstiziale calcificata. aDNA è dunque recuperabile da ossa e denti, resti mummificati naturalmente o artificialmente, manufatti, coproliti, campioni inglobati in ambra, preparati istologici paraffinati, campioni secchi o disidratati, conservati in ambienti aridi o musei, campioni umidi (da torbiere) o congelati (da ghiacciai, tundre).
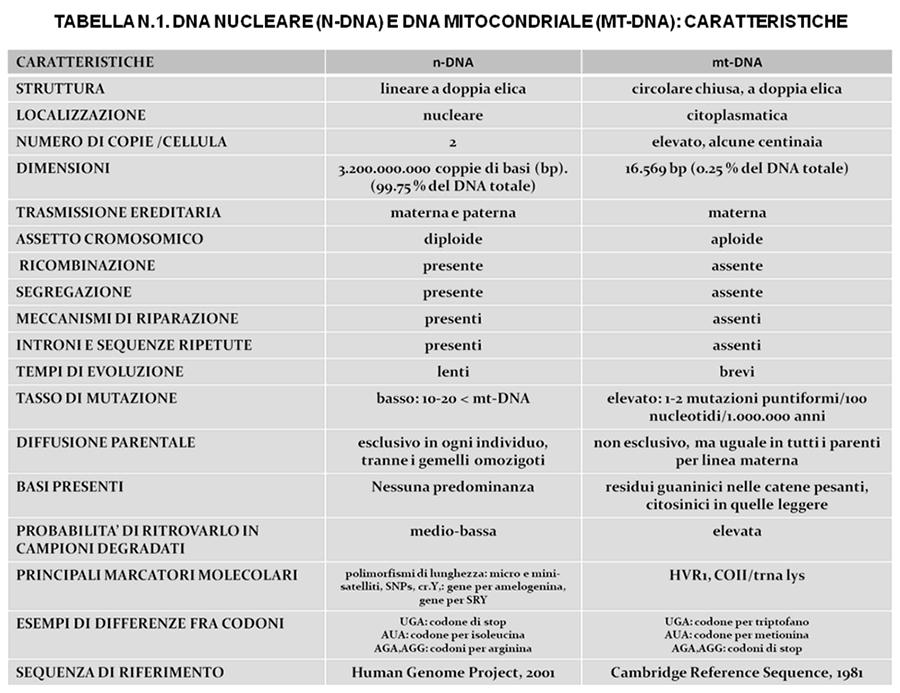
Il DNA mitocondriale (mtDNA), nello studio dei genomi antichi, si presta meglio di quello nucleare (nDNA) per diversi motivi7: in una cellula è presente in numerose copie, le probabilità di reperirlo non danneggiato sono elevate, è molto fedele nella ricostruzione delle linee evolutive in quanto trasmesso solo per via materna, ha un tasso di evoluzione elevato, non è sottoposto a eventi di ricombinazione e segregazione. Nella tabella n. 1 sono illustrate le differenze fra n/mtDNA.
mtDNA e, in parte, nDNA antico sono stati esaminati, tra gli ominidi estinti, in h.neanderthalensis, uomo di Denisova e confrontato con quello di h.sapiens di varie epoche; i risultati migliori sono stati ottenuti con reperti di un’epoca comprea fra 60.000 e 20.000 years ago (ya), comunque a tutt’oggi non è mai stato estratto materiale genetico su fossili più antichi di 100.000 ya. H. sapiens ha convissuto in Europa ed Asia con queste specie, fino all’estinzione di queste ultime, scambiandosi frammenti genetici di cui l’uomo moderno porta ancora l’impronta nel proprio DNA. L’uomo di Denisova è la prima specie, nel genere “homo”, identificata mediate esami molecolari sul DNA antico.
I principali fattori che condizionano la presenza o meno di aDNA in un campione e la sua degradazione, sono ambientali e relativi al sito di conservazione: temperatura, umidità relativa, pH, esposizione ai raggi solari, composizione in molecole organiche e inorganiche, quadro microbiologico; habitat caldo-umidi, con molta acqua, ossigenati favoriscono la crescita di microrganismi i cui enzimi denaturano senz'altro gli acidi nucleici; ambienti aridi con caldo secco (deserti), freddo e circolazione di aria gelata (steppe siberiane), scarsa ossigenazione e alta salinità offrono invece condizioni opposte e ottimali alla conservazione. Il termine tecnico che riassume le modificazioni chimiche cui va incontro il DNA, senza i fisiologici meccanismi riparativi di una cellula vivente, è “diagenesi". Le conseguenze sono quantitative e qualitative: fra le prime, la frammentazione della molecola. Le principali alterazioni chimiche consistono in: depurinazione, idrolisi dei legami fosfodiesterici, azione di radicali liberi, ossidazioni, deaminazione, dimerizzazione delle pirimidine, reazioni di alchilazione.
Contemporaneamente alla degradazione, si verifica quasi inevitabilmente un altro evento che aumenta le problematiche relative all'analisi di aDNA: la contaminazione del reperto che riconosce diverse cause: 1) contaminazione biologica, ad opera di microrganismi ambientali, batteri, miceti, protozoi, che sostituiscono il genoma del reperto col proprio in percentuali variabili a seconda dell'età e della carica microbica accumulatasi col tempo. 2) contaminazione da parte degli operatori, sia di un elemento di nuova individuazione, che già studiato e musealizzato; quanto avvenuto in passato non può più essere cancellato, oggi esistono linee-guida che riguardano reperti vecchi e nuovi in modo da formare barriere tra questi e le diverse figure di tecnici, archeologi, studiosi di varie discipline che vengono in contatto diretto con l'oggetto di studio. 3) contaminazione attraverso apparecchiature e reagenti di laboratorio: la prevenzione di possibili inquinamenti richiede la sterilità assoluta di macchinari, vetreria, soluzioni, reagenti; l'impiego di cappe sterili; l'adozione di barriere fra operatori da una parte, reperti, macchinari e reattivi dall'altra; la costruzione di laboratori specifici ove siano ben separati gli ambienti dedicati alla fase pre-amplificazione (prelievi, estrazione), dall'amplificazione stessa alle fasi post-amplificazione (clonaggio, sequenziamento).
E' il tempo il fattore determinante che condiziona la presenza o meno, la parziale integrità o la frammentazione irreversibile fino alla disgregazione del materiale genetico in un reperto antico; le condizioni di conservazione, i diversi elementi chimici, fisici e biologici sono senz'altro importanti, ma il periodo trascorso fra la fine di una vita e il recupero di quanto fisicamente rimasto rappresenta l'ostacolo principale alla possibilità di condurre indagini su aDNA.
Dobbiamo affidarci a prove indirette per stimare la reale possibilità di recuperare materiale genetico antico: il rapporto di racemizzazione dell'ac. aspartico è un marker affidabile in quanto correla col principale meccanismo degradativo di aDNA: la depurinazione. Se il rapporto D/L Asp è = o < a 0.1, si può recuperare aDNA e procedere con l'estrazione e l'amplificazione; se il rapporto è > a 0.1 le possibilità di trovare aDNA sono minime o nulle e dunque la procedura dovrebbe essere interrotta. La termogravimetria sfrutta la proprietà di un campione, come ad es. un osso, di perdere componenti differenziate se sottoposto all'azione del calore a diverse temperature: fra 200 e 600° si perdono le componenti organiche, tra cui gli ac.nucleici. In "Science 2000; 289: 1139-54" sono illustrate le linee-guida per esaminare aDNA e risolvere i problemi brevemente esposti.
I principali marcatori genetici utilizzati sono: marcatori mitocondriali, high variability region I (HVRI), HVRII e HVRIII: indicati nei confronti fra popolazioni separatesi durante l’evoluzione, ricostruzione di antiche migrazioni e colonizzazioni, identificazioni individuali in una linea familiare e linee di parentela; regione di controllo del Citocromo C Ossidasi subunità II (COII/trna lys): studi a livello di popolazioni. Marcatori cromosomi sessuali: gene per amelogenina (dimorfismo di lunghezza fra X e Y), Sex determining region Y (SRY), Short Tandem Repeat (STR o microsatelliti): determinazione del sesso, genetica di popolazioni e ricostruzione dei processi migratori. Marcatori nucleari: polimorfismi di lunghezza: micro e mini-satelliti, polimorfismi a singolo nucleotide (SNPs): tipizzazione genetica di un individuo, ricostruzione legami genealogici fra gruppi di individui, confronto fra specie coeve o lontane nel tempo, storia evolutiva di una specie.
Primi dati acquisiti.
Il numero di differenze genetiche nelle sequenze di mtDNA che separa due popolazioni è direttamente proporzionale al tempo a partire dal quale esse hanno condiviso la stessa antenata materna: quanto più a lungo sono state separate, tanto maggiore sarà l’accumulo di mutazioni casuali con cui i geni si evolvono; questo accumulo è definito tasso di evoluzione: per mtDNA è di 1-2 mutazioni puntiformi/100 nucleotidi/milione di anni: molto elevato e superiore di 10-20 volte quello di nDNA. mtDNA viene assunto come un “orologio molecolare” che batte con un ticchettio molto veloce e costante nel tempo, in termini di evoluzione, e che si presta a documentare gli studi di antropologia molecolare: l’evoluzione di specie che si sono diversificate o di popolazioni umane che si sono differenziate. Il termine di orologio molecolare fu adottato nel 1962 da E.Zuckerland e L.Pauling, che si avvalsero delle mutazioni riguardanti l’emoglobina; successivamente vennero utilizzate altre proteine come le immunoglobuline, infine i marcatori genetici. La rappresentazione grafica di questi dati permette la costruzione di alberi genealogici evolutivi.
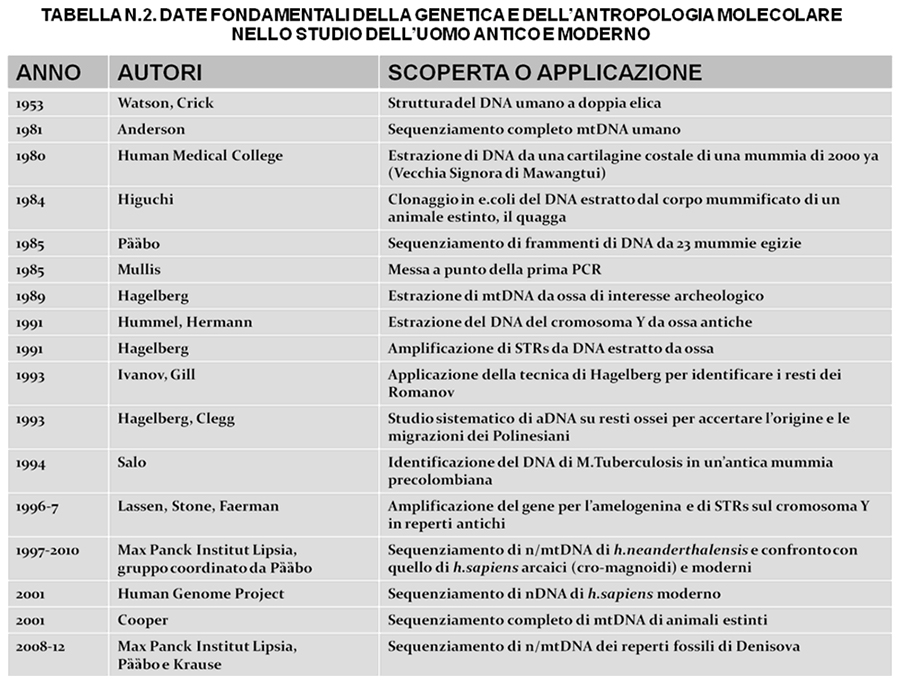
La linea di discendenza mitocondriale è dovuta ad un antenato comune collocato alla radice dell’albero filogenetico che ha trasmesso il suo mtDNA inalterato, se non per mutazioni casuali. E’ stato dimostrato che tutti i tipi di mtDNA presenti attualmente nella nostra specie sono derivati da un’antenata comune che visse in Africa circa 200.000 ya: da qui è nata la teoria dell' "Eva mitocondriale“ che conferma la nostra origine monocentrica, africana e abbastanza recente da un membro del "gruppo fondatore umano", che ha trasmesso alla discendenza il proprio mtDNA sostanzialmente invariato; il che non esclude che vi fossero altre donne con altri tipi di mtDNA, ma queste non hanno potuto trasmettere i propri caratteri presumibilmente perchè si sono estinte, come molte prove archeologiche e paleontologiche hanno di recente confermato, che h. sapiens attuale deriva da un piccolo gruppo africano che sopravvisse a catastrofi demografiche conseguenti a cause alimentari e climatiche. Nella tabella n.2 abbiamo riportato alcune date fondamentali nello studio molecolare e genetico della specie umana, antica e moderna.
Dati recenti e prospettive.
Il genoma è non solo contenitore di tutta l'informazione genetica, ma museo esso stesso del passato dell'uomo, delle specie ancestrali, un museo che, imparando a percorrere e a decodificare, può rivelare tantissimo della nostra storia biologica e mentale. Concludiamo presentando alcuni risultati recenti, mediante analisi molecolare e genetica, in diversi campi dell'evoluzione umana.
1) Ricostruzione della storia evolutiva della nostra specie. Confronto con i genomi di altri “homo” e di primati. Dati paleoantropologici e genetici concordano nel riconoscere le prime tracce di h.sapiens 200.000 ya in Africa: il suo antenato più prossimo fu probabilmente h.heidelbergensis, da cui si era staccato almeno 300.000 anni prima h.neanderthalensis. Dopo aver più volte rischiato l’estinzione per crisi demografiche e alimentari, la nostra specie cominciò a lasciare l’Africa circa 80.000 ya e raggiunse l’Eurasia 45.000 ya, che trovò già occupata da almeno altre due specie di homo: il già citato neandertaliano e il misterioso “Uomo di Denisova”, entrambi destinati ad estinguersi3. Prima di rimanere totalmente solo, h.sapiens si è sicuramente incrociato con le altre specie, scambiandosi frammenti genetici di cui oggi rimangono tracce nel nostro genoma. Confrontando i genomi, è stato dimostrato che H. Sapiens condivide con i primati almeno il 90 % del DNA; il 10 % divergente riguarda soprattutto l’espressione funzionale dei geni e meccanismi di trascrizione-traduzione. Per oltre il 30 % il genoma umano è più prossimo a quello dei gorilla che degli scimpanzè, soprattutto nei geni che regolano lo sviluppo cerebrale e l’udito. Con i primati e le altre specie homo con cui abbiamo convissuto condividiamo le basi genetiche del sistema ABO. Abbiamo ereditato il 3-4 % del genoma da h. neanderthalensis5 e il 5-6 % dai denisoviani. Gli incroci genetici hanno arricchito quali-quantitativamente il nostro sistema immunitario e il numero di antigeni HLA: HLAB73 deriva dai Denisoviani, HLA11 dai neandertaliani asiatici. La nostra specie ha almeno 200 regioni genomiche specifiche che si sono evolute diversamente dalle altre specie e deputate alla regolazione di processi metabolici; abbiamo 510 regioni regolatorie che mancano alle altre specie ominine, ma abbiamo anche perso geni come Gadd45d la cui assenza sblocca l’implementazione di funzioni cognitive superiori. Homo sapiens e neanderthalensis hanno volumi cerebrali quasi simili, ma cambia l’organizzazione delle aree cerebrali, soprattutto quelle pre-frontali. Dati archeologici, antropologici e genetici confermano nel porre la divaricazione ultima uomo scimmia 6.500.000/7.000.000 ya: nessun fossile anteriore, a tutt’oggi, può essere inserito tra gli antenati o precursori ominidi. L’origine di un’andatura bipede “moderna” è fatta risalire a 4/4.500.000 ya, ma fossili anteriore recentemente scoperti e allo studio potrebbero retrodatare l’acquisizione di questa fondamentale caratteristica umana.
2) Percorsi geografici di h.sapiens: migrazioni e colonizzazione della terra. Pur iniziando ad affacciarsi “Out of Africa” oltre 100.000 ya, h.sapiens ha sistematicamente colonizzato il mondo a partire da 80.000 ya fino a raggiungere la Terra del Fuoco 13.000 ya. Calcolando la distanza fra Etiopia e punta sud del Cile, 25.000 Km, sono stati necessari 67.000 anni e 2.300 generazioni. La colonizzazione del pianeta è studiata in rapporto alla diffusione e alla frequenza degli aplogruppi mitocondriali e al tasso di mutazione genica: ad esempio, il popolamento del continente americano, favorito dall’unione via terra con l’ Asia, da parte di cacciatori siberiani , di cui rintracciamo gli aplotipi ABCDQ nei nativi americani, iniziò 25.000 ya e si concluse 13.000 ya, seguendo una rotta terrestre ed una costiera9. Fu un’impresa epica, così definita dall’archeologo Bordes: “Il popolamento del nuovo mondo, dal freddo assassino dell’ artico, all’umidità soffocante dell’ Amazzonia, ai venti di tempesta della Terra del Fuoco, è una delle più grandi imprese dell’umanità, è una storia di resistenza e adattamento che rimarrà senza eguali fino a quando l’uomo non atterrerà su un pianeta di un’altra stella”. Nel loro lungo percorso nord-sud attraverso sterminate praterie, montagne e foreste del continente americano e lungo le coste del pacifico, i colonizzatori preistorici hanno lasciato numerose tracce archeologiche, manufatti che ancora oggi riemergono ovunque.
3) Geni e fisiologia: l'esempio del linguaggio. Per acquisire questa nuova funzione nella nostra specie, segno di diverse e peculiari manifestazioni cognitive, si sono rese indispensabili: a)modificazioni anatomiche, b)coordinazione neuro-muscolare, c)sviluppo di connessioni e circuiti cerebrali, d)modificazioni e attivazioni geniche: per a) e d) possiamo recuperare tracce antropologiche e molecolari. Le modificazioni anatomiche acquisite con l'evoluzione si apprezzano dallo studio e dal raffronto di fossili di antichi ominidi, primati, h.sapiens arcaico e moderno e ricostruzioni computerizzate del tratto vocale. A livello molecolare, importante è il gene Foxp2: situato sul braccio lungo del cromosoma 7, codifica per un fattore di trascrizione importante nello sviluppo del cervello, del polmone, di strutture neurali connesse con il controllo motorio e l'articolazione del linguaggio. Mutazioni del gene sono associate a dislessia, disprassia e disturbi cognitivi. Ritenuto specifico della nostra specie (è assente in tutti i primati), nel 2007 è stato trovato a sorpresa nel genoma di h.neanderthalensis (che poteva articolare solo pochi suoni), rimettendo in discussione tutte le nostre conoscenze su origine ed evoluzione del linguaggio10.
4) Coevoluzione geni-cultura. L’esempio della lattasi. " In passato l’innovazione culturale ha spesso diretto l’evoluzione biologica; la coevoluzione biologica e culturale ha permesso all’uomo di predominare anche su specie più forti fisicamente; ciò fu possibile sia grazie alla maggior intelligenza che all’accumulo di conoscenze, trasmissibile fra le generazioni, che chiamiamo “cultura” (L.L.Cavalli-Sforza). La persistenza del gene per la lattasi ne è un esempio4: si dovrebbe disattivare progressivamente dopo lo svezzamento, ma negli ultimi 10 millenni, in coincidenza con la rivoluzione neolitica, l’inizio dell’agricoltura e la domesticazione di piante e animali, si sono selezionate varianti geniche (per un polimorfismo a singolo nucleotide o SNP) che codificano enzimi persistenti anche nell’adulto, che può così digerire a lungo o per sempre il latte. La mutazione genetica è coincisa con l’acquisizione delle tecniche di allevamento e domesticazione, dal medio-oriente e poi ad altri paesi, di animali fornitori di latte. Il gene è mutato con maggior frequenza in popolazioni che hanno, in tempi brevi, acquisito livelli culturali e tecnologici nell’allevamento più avanzati, frequenza bassa o nulla nella situazione opposta.
![]()
![]() q0 S
= coefficiente di selezione
q0 S
= coefficiente di selezione
log ______ T = tempo espresso in generazioni
1 - q0 Q0 = frequenza genica
S = ___________ 1 - q0 = differenze di frequenze
T
Questa formula permette di calcolare, per un gene, il coefficiente di selezione S8; per il gene della lattasi il valore ottenuto varia da 1.5 – 3 per tempi lunghi in popolazioni con scarso sviluppo tecnologico e culturale, a 10 (il più elevato registrato per un gene) per tempi brevi e tecnologia avanzata.
5) Malattie umane antiche e nuove. Evoluzione e cambiamenti nelle interazioni fra patogeni ed ospite. L'esempio della tubercolosi. Nel 2002, in una cava di travertino nei pressi della città di Kocabas, Anatolia, sono riemersi i resti di un cranio di un giovane maschio di h. erectus di 500.000 ya. Le indagini radiologiche in 3D, mediante scansione con radiazione di sincrotrone e quelle molecolari hanno rivelato come egli fosse affetto da leptomeningite tubercolare: questa eccezionale scoperta ha permesso di retrodatare le più antiche prove di tale malattia, prima accertate su alcune ben più recenti mummie egizie e peruviane. 10.000 anni fa nacquero i primi agglomerati urbani: l'affollamento, la scarsa igiene, il contatto con animali vettori comportò la diffusione di malattie infettive quali vaiolo, tubercolosi e lebbra; ma contemporaneamente si sono verificati cambiamenti adattativi che hanno determinato una selezione di individui geneticamente resistenti. Correlando mutazioni vantaggiose che conferiscono resistenza alla tubercolosi e alla lebbra (ad esempio la presenza dell'allele SLC11A1-1729-55del4) con l'epoca di fondazione delle città, Barnes e coll.2 hanno dimostrato che la frequenza di queste mutazioni è tanto più diffusa quanto maggiore è l'antichità dell'agglomerato urbano; nelle città fondate di recente e nelle campagne la mutazione vantaggiosa è rara. In pratica nelle città ove il rischio di contagio era alto, si è verificata una selezione: è un esempio di evoluzione dinamica, in divenire, che rimarca aspetti recenti dell'evoluzione umana, in particolare l'importanza delle città e delle aggregazioni come forza selettiva; questi meccanismi spiegano le differenze di frequenza nella resistenza alle malattie infettive che registriamo nel mondo.
6) Ricostruzione di antiche vicende storiche. Diverse le vicende storiche che l'indagine molecolare ha aiutato a interpretare: ricordiamo l'Identificazione individuale e le relazioni parentali ricostruite dai resti ossei dei Romanov, l' attribuzione di sesso, età biologica e antichità date alle ossa trovate nella tomba di Petrarca, ma soprattutto l'indagine dei rapporti parentali e delle cause morbose che portarono all' estinzione della XVIII dinastia faraonica1; abbiamo pertanto conferma della ripetuta pratica del matrimonio tra congiunti, di quali fossero genitori e sposi di ogni membro della famiglia, di come Tutankhamun e la sposa-sorella avessero generato solo feti morti; ma soprattutto è emerso il complesso quadro patologico che ha indotto a parlare di "crepuscolo genetico" della dinastia e di uno dei periodi più oscuri della storia egiziana. Le indagini molecolari e antropologiche condotte sulle mummie reali hanno rivelato che Tutankhamun non morì per cause traumatiche, che era affetto da malaria e diverse patologie congenite: M. di Köhler II, sindrome di Freiberg-Köhler con necrosi settica ossea che determinarono iposviluppo e, verosimilmente, immuno-deficit. Tutte queste patologie sono state ritrovate nella maggioranza delle mummie reali. Ritenendosi divini e icontaminabili, gli ultimi rappresentanti della dinastia si unirono solo fra loro trasmettendosi diverse patologie congenite e non e divenendo incapaci di generare una progenie sana.
7) Ricostruzione di antiche usanze e percorsi culturali. Riportiamo un antico caso di “pietas” alle porte di Roma. Necropoli di Caracupa, Sermoneta (LT), IX-VIII secolo a.c., area nota come “Latium vetus”; tomba n.12, giovane donna, aa 25/27, di alto lignaggio, con ricco corredo funebre. La ricognizione antropologica e poi genetica ha evidenziato una grossa massa cerebrale, ipotrofia delle gambe, scarsi segni di inserzioni muscolari; era affetta da una rara variante genetica di “acondroplasia congenita” che provocò ritardo mentale e incapacità alla deambulazione12. La disabile crebbe fino ad un’età adulta in una comunità benestante e fu sepolta con oggetti tipici del lavoro femminile: tessitura e cucina, ma volutamente spezzati, a indicare l’impossibilità a usarli da parte dell’inumata. La singolarità del caso è da interpretare come un omaggio alla defunta disabile, un messaggio per i posteri e rappresenta secondo noi un caso di “pietas” e una lezione di civiltà sociale, ben 3000 anni fa.
Testimonianze archeologiche e antropologiche riportano riti funerari simili a quelli di Caracupa nella Troade, anche più antichi; le affinità genetiche riscontrate ieri e oggi fra uomini e animali addomesticati di quella regione anatolica, di aree del Lazio e dell’antica Etruria, potrebbero su nuove basi rinvigorire uno dei miti classici più noti ma controversi, nato in verità in epoca arcaica e ripreso per scopi propagandistici in età augustea: quello del pio Enea e la leggenda della fondazione di Roma ad opera di discendenti degli esuli troiani. “Dietro ogni mito c’è sempre, in percentuale variabile, un po’ di verità storica”.
Conclusione. Le nuove tecnologie molecolari rappresentano una formidabile occasione per implementare lo studio del passato umano, antico e recente, affiancandosi alle indagini convenzionali antropologiche, archeologiche e paleontologiche. I primi risultati, di cui abbiamo riportato alcuni esempi significativi, sono al tempo stesso esaltanti e incoraggianti. Ulteriori progressi arriveranno con la miglior conoscenza del genoma umano, sia moderno che antico. “La nostra storia evolutiva (e la sua conoscenza) è essenziale per comprendere le vicende storiche del passato dell’umanità, con le varie rivoluzioni biologiche, socioeconomiche e culturali che l’hanno attraversato, ma anche il nostro presente che è già futuro: il futuro dell’informazione globale e delle innovazioni tecnologiche, è radice di tutto questo e delle prossime evoluzioni umane”11.
BIBLIOGRAFIA ESSENZIALE
1) Awass Z et al. Ancestry and pathology in king Tutankhamun’s family. JAMA 2010; 303 (7): 638-47
2) Barnes I,
Duda A, Pybus OG et al. Anciet urbanization predicts genetic resistance to
tuberculosis.
Evolution 2011;
65(3): 842-8
3) Belli F. Conoscenze biomediche convenzionali e indagini di laboratorio innovative in paleoantropologia. Il Policlinico 2011; 118 (1): 1-22
4) Beja-Pereira A, Luikart G, England PR et al. Gene-culture coevolution between cattle milk protein genes and human lactase genes. Nature Genetics 2003; 35: 311-3
5) Burbano HA, Hodges E, Paabo S. Targeted investigation of the Neandertal genome by array-based sequence capture. Science 2010; 328: 723-5
6) Caramelli D. Antropologia molecolare. Firenze Unniversity Press ed, Firenze, 2009
7) Caramelli D, Lari M. Il DNA antico. Metodi di analisi e applicazioni. Pontecorboli ed, Firenze, 2010
8) Cavalli-Sforza LL, Menozzi P, Piazza A. Storia e geografia dei geni umani. Adelphi ed, Milano, 1997
9) Erlandson JM. Paleoindian seafaring, maritime technologies and coastal foraging on California's channel islands. Science 2011; 331: 1181-5
10) Krause J, Lalueza-Fox C, Orlando L et al. The derived FOXP2 variant of modern humans was shared with Neandertals. Curr Biol 2007; 17 (21): 1908-12
11) Ortino S. La struttura delle rivoluzioni economiche. Cacucci ed, Bari, 2009
12) Rubini M. Una nuova tomba femminile da Caracupa Sermoneta. Nota antropologica. In: Rizzo S (a cura di), Roma città del Lazio, Catalogo della mostra, Roma, 2002